

日本のヒグマとツキノワグマの個体数は、過去数十年間で著しく増加しました。2025年には、ほぼ1日に1件の割合で市街地や農地でのクマの出現が確認され、そのニュースは世界各地で報じられました。本稿では、クマの空間的個体群動態、クマと人間の遭遇、クマの個体群管理の考え方について考察していきます。本稿の目的は、生態学者ではない読者に空間的個体群制御モデルへの理解を深めていただくことです。また、本稿ではクマを例として議論を展開しますが、その理論の多くは大型肉食動物の個体群動態モデリング全般に適用可能です。
遭遇・襲撃・死亡事例:日本とスカンジナビアの共通点
スカンジナビアヒグマ研究プロジェクトの観察データをもとに、日本とスカンジナビアにおけるクマ襲撃事例について興味深い比較をすることができます。両地域の状況は大きく異なる一方で、共通点もあります。
1970年から2016年にかけて、スカンジナビアでは44件のクマによる人への襲撃が発生し、うち2件が死亡事故です。この数字だけに注目すると、2025年時点でほぼ毎日クマの襲撃が発生し、多数の死亡者が出ている日本とは全く異なる状況にあります。では、共通点はどこにあるのでしょうか。
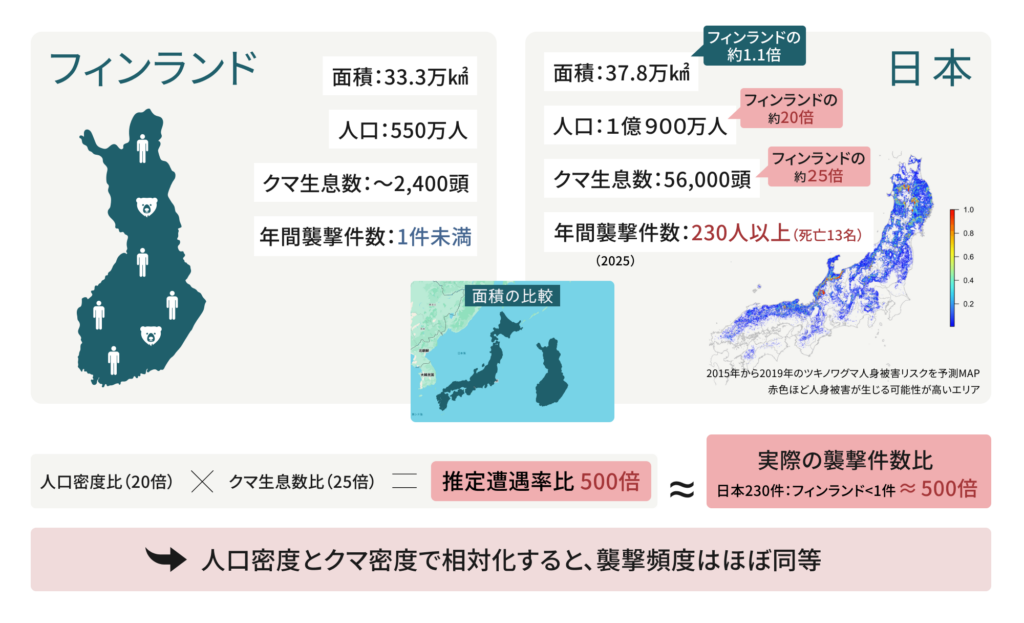
注目すべきは、クマと人間の遭遇率です。例えば、スカンジナビア地域の一国で、Think Natureの最高科学責任者(CSO)の母国でもあるフィンランドを例に挙げて考えてみます。フィンランドの人口は550万人であり、面積は北海道と本州を合わせたものとほぼ同等です。一方で、北海道と本州の人口は1億900万人であるため、日本とフィンランドの人口密度の比率は20倍となります。また、最新のデータによると、フィンランドのクマ生息数は2,000~2,400頭であるのに対し、日本は56,000頭であり、その比率は25倍となります。そして、フィンランドでのクマによる襲撃は年間1件未満であるのに対し、2025年に日本で発生したクマによる襲撃は235件であり、死亡者は13名でした(2025年12月10日 NHK報道)。両国の人とクマの密度を組み合わせると、人とクマの遭遇率比(人口密度比(20倍)×クマ生息数比(25倍))は500倍と推定することができ、これは実際の一年あたりのクマ襲撃件数の比率と近い値となります(日本:253件、フィンランド:<1件)。したがって、スカンジナビアと日本におけるクマ襲撃件数の絶対値は大きく異なるものの、遭遇の基盤となる人口密度とクマ密度で相対化すると、襲撃頻度はほぼ同等であることがわかります。
さらに、スカンジナビアにおけるクマの襲撃件数に対する死亡者の比率は4.5%(2/44)であり、この値は2025年に日本で観測された5.5%(13/235)に非常に近い値となります。したがって、クマ襲撃の約5%が死亡事例を引き起こすと考えられます。
大型哺乳類の空間個体群モデル構成要素
スカンジナビアヒグマ研究プロジェクト(SBBRP)は、40年以上にわたる実証的研究に基づいて350本の学術論文を発表するなど、ヒグマに関する重要な研究を実施してきました。ここでは、クマ個体群管理のモデリングを検討する前に、SBBRPによる研究成果を集約した40年報告書(Tallian et al. 2017; https://www.brownbearproject.com/post/new-reports-40-years-of-reseach-by-the-sbbrp)に記載されたクマの基本的な生態学的特徴を紹介します。なお、スカンジナビアのヒグマは生物学的に北海道の個体群と極めて近縁であるため、多くの生態学的特性は本質的に同一であると考えられます。
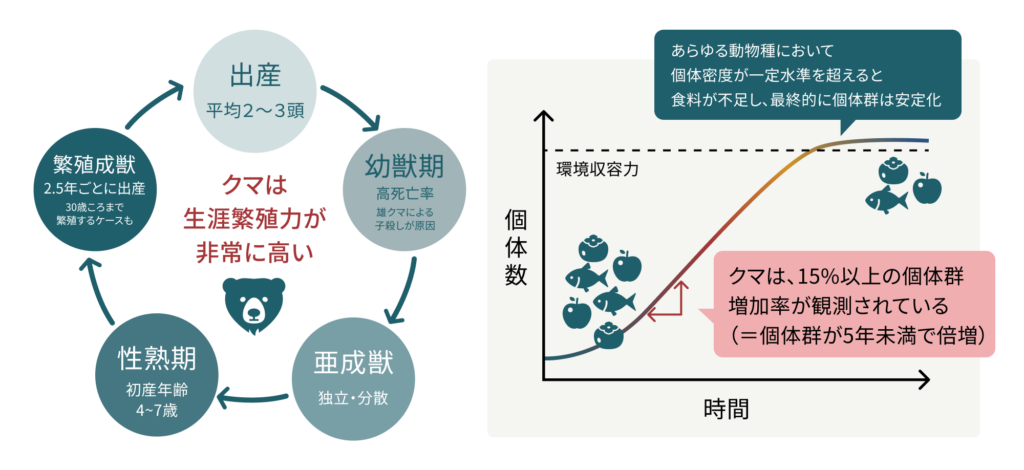
死亡率(あるいは生存率)と繁殖(出産と死亡)は、個体群モデルの基礎的な要素です。ヒグマの性成熟年齢は4~6歳であり、性別によって異なります。出産数は、通常2~3頭です。幼獣の死亡率は高く、血縁関係のないオスによる幼獣殺害が主な原因となります。メスは平均2.5年ごとに出産し、25歳を超えても繁殖能力を維持するため、生涯繁殖力は非常に高く、野生では30歳を超えて繁殖する個体も確認されています。
個体群の増加は、出産数と死亡数の差によって生じます。個体群増加率とは、資源が豊富で狩猟の影響がない状態における個体群の増加率のことです。クマでは15%以上の固有増加率が観測されており、これは個体群が5年未満で倍増することを意味しています。なお、あらゆる動物種において、個体密度が一定水準を超えると食料が不足し、最終的に個体群は安定化します。
SBBRPがヒグマの高い固有増加率を報告するだけでなく、日本からの情報もまたツキノワグマの非常に高い個体増加率を裏付けている。2012年の日本のツキノワグマの個体数は約15,000頭と推定され、10年間で約44,000頭まで増加した。10年間で個体数が3倍になったということは、増加率が11~12%であることを示しています。
資源は、死亡率や固有増加率に続き、空間個体群モデルの第二の主要要素になります。クマは雑食性・捕食性・腐食性を併せ持ち、多様な食物資源に依存しています。例えば、ベリー類、ドングリ、木の実、草本・顕花植物、菌類、有蹄動物(シカ、イノシシなど)、アリ、魚類などを食べます。加えて、食物資源の利用は季節や年次、個体群によって変化します。そのため、クマの年間移動は食料の入手可能性によって部分的に説明することができ、これは人間とクマの遭遇リスク区域を予測する際に重要となる可能性があります。実際に、クマの主要な食料源に関する種分布モデルは、年間サイクルを通じてクマがどこに集中するのかを知るうえで大きな助けとなります。また、資源という観点では、クマは、食料だけでなく冬眠用の穴も必要としています。
クマを含む多くの動物は、採食や繁殖に利用する行動圏を持ちます。スカンジナビアでは、成体のオスの行動圏が最も大きく、資源の入手性や季節によって500㎢~1,000㎢の規模になります。単独行動のメスの行動圏ははるかに小さく、150㎢~250㎢です。そして、子連れのメスの行動圏が最も小さく、おそらく50㎢程度とされています。これは、見知らぬオスとの接触を避けるためです。なお、クマの行動圏は互いに重なり合っています。
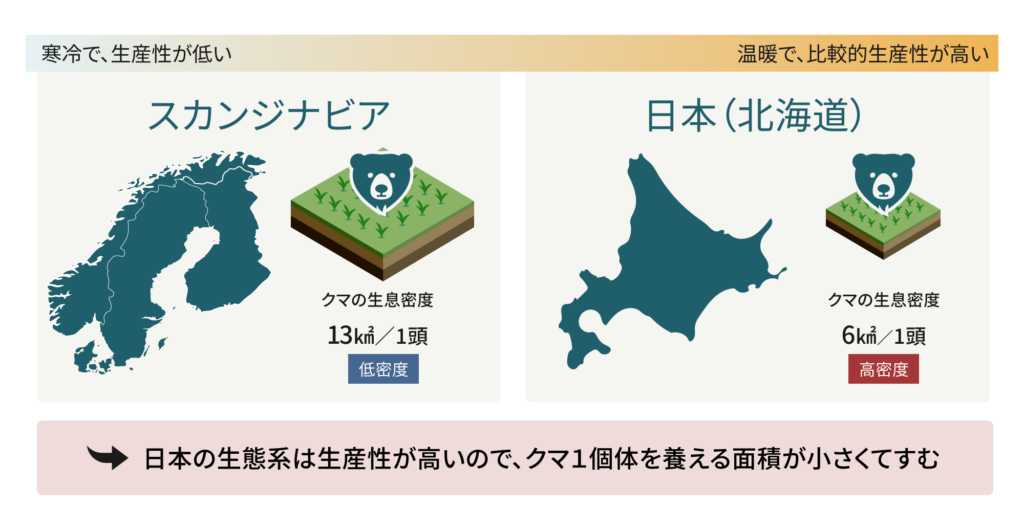
クマの生息密度は資源密度に大きく依存します。スカンジナビアでは、観測された最大のクマの生息密度は13㎢あたり1頭です。温暖な中央ヨーロッパのスロベニアでは、最も適した地域で0.5~3.5頭/㎢に達します。そして、日本のクマの密度は、スカンジナビアよりも高いと考えられます。これは、日本の島々は一般的に生産性が高いためです。北の島である北海道でさえ、近年は13,000頭以上のヒグマが生息し、これは6㎢あたり1頭に相当します。このような、人里から離れた主要生息地域における密度推定値は、人間が居住する地域に近い場所でのクマの個体群増加の可能性を予測する際の助けとなります。
空間個体群モデルの第三の主要要素は、生息地の利用と移動(分散・回遊)です。クマの生息地利用は、主に餌の入手性と視覚的な遮蔽物(隠れ場所)の好みによって決まります。クマは、森林や険しい地形を好む傾向がありますが、特に夕暮れ時や暗闇ではより開けた地域も利用します。特に、オスのクマは家屋、林道、林業施設などの人間の存在を示す痕跡がある地域を避ける傾向にあります。一方で、子連れのメスは、幼獣を殺すオスを避けるため、人間の居住地に比較的近い場所に留まることがあります。また、食料不足や狩猟の不在に起因する慣れが、人間に対する回避行動を鈍らせる場合もあります。加えて、クマの生息地利用は季節や時間帯によって変化するため、クマへの遭遇リスクにも同様の変動が生じます。
行動圏の規模推定とクマの移動情報は、個体群動態に関連する空間スケールの理解に役立ちます。クマは長距離を移動することで知られ、最も長く移動するオスは、スカンジナビアで最大470kmの移動をした事例が報告されています。一方で、子連れのメスの移動は最も短く、スカンジナビアの事例ではメスの最大移動距離は90kmです。多くの若齢のクマは移動・分散して新しい場所へと定着します。また、局所的な個体群密度が高いほど、若齢のクマの分散発生確率が増加します。分散距離は容易に数十kmに達するため、個体数が増加するとクマの個体群は比較的急速に拡大する傾向があります。なお、クマは泳ぎが得意であり、比較的急な斜面も移動できるため、分散における障壁は少ないです。つまりは、クマが行く「能力」よりも、そこへ行きたい「意思」の方が重要なのです。
クマの個体群動態に関する特筆すべき観察結果
クマと人間の衝突を考える際、以下の観察結果が関連性を持つと考えられます。クマは雑食性であること。非常に高い個体群増加率を示すため、個体群密度が低い場合、その個体群が倍の数になるまでに5年もかからないこと。日本のクマの個体数は非常に多く、年間数千個体が誕生しており、クマ個体群は保全生物学的な脅威を全く受けていないこと。近交弱勢の悪影響を受けないとされる最低限の個体数である1,000と比較して、日本のクマ個体群はその数倍の規模にあること。
非常に長い距離を移動できることも重要な観察結果の一つです。これは、急速な個体数の増加と相まって、食物や生息地が十分に存在するすべての地域へとクマの個体群が拡大し、やがて定着することを意味しています。クマには人間以外の天敵が存在しません。生息地の喪失とそれに伴う資源の不足が、クマが恒久的な縄張りを確立できる場所を制限しているのです。
日本におけるクマによる襲撃事件の多発は、クマ個体群が既に遠隔地まで拡大し、個体数増加がクマを人間の居住地域に近づけていることを実証的に示しています。では、クマの襲撃を理解し、クマの個体群を管理するためには、どのようなアプローチが考えられるのでしょうか。
メカニスティックモデルと現象論的モデル
メカニスティックモデルと現象論的モデルには大きな違いがあります。医学を例に考えてみます。喫煙が多くの疾患の発症率を著しく高めることは知られており、大規模な実証データからも観察できます。この現象を観察するために、当該疾患がどのように発症するのかというメカニズムを解明するための生化学的・生理学的モデルは必要ありません。禁煙がリスクを低減することを理解するために深い分析は必要ないのです。
クマについても同様の状況が存在します。日本では長年にわたるクマの襲撃地に関する豊富なデータが存在し、これらのデータに基づき危険地域が特定されています。そのため、クマとクマが利用する資源に関する完全なメカニスティックな空間個体群モデルを開発せずとも、クマ襲撃という現象をほとんど理解することができます。
一方で、メカニスティックモデルの利点として、景観動態や順応的な個体群管理の具体的手法など、任意の生態学的知見を組み込めることが挙げられます。このモデルは、「なぜ」を解明することに優れ、理解を深めたり仮説を立てたりすることに向いています。しかし、重大な欠点も存在します。メカニスティックモデルの複雑性は、モデルの構築やパラメータ推定、検証に多大な労力を要することを意味しています。実際、メカニスティックな空間個体群モデルは、全モデルパラメータの統一的な統計的推定をおこなうには過度に複雑です。例えば、パラメータ間の相関関係が無視された場合、誤った予測が出力される可能性があります。
移動と分散の現実的なモデリングは著しく困難であり、偶発的に複雑化されることもあります。移動と分散のメカニスティックモデルに関連する生物学的要素には、以下の7つが含まれます。(1)生息地選好性、(2)生息地間の遷移確率、(3)異なる生息地における移動速度と死亡率、(4)境界追従行動、(5)相関付きランダムウォークの種固有パラメータ、(6)個体間変動、(7)他種との相互作用。
現象論的モデルは「なぜ」ではなく「何」に焦点を当てます。このモデルは、統計学や機械学習の手法を用いて、観察されたパターンと説明変数を適合させることに優れています。一方で、現象論的モデルの限界として、個体群動態に関する生態学的仮説の生成と検証には有用性が限定されているという点が挙げられます。なお、クマへの遭遇の現象論的モデルでは、人間とクマの個体群密度分布図が主要な構成要素となります。
メカニスティックモデルや現象論的モデル以外にも、個体群生物学で頻繁に用いられるモデルとして、構造化個体群モデルとメタ個体群モデルがあります。構造化個体群モデルは、漁業で一般的に使用されている単一個体群モデルであり、通常は年齢(生活史段階)と性別によって構造化された個体群サイズをモデル化します。しかし、構造化個体群モデルは、クマのモデリングには理想的でありません。なぜなら、クマによる襲撃の問題には、空間的要素が存在するためです。一方で、メタ個体群モデルでは、主要な簡略化の手段として、(1)環境を生息地と非生息地に分ける二分法と、(2)局所的な動態を絶滅-定着プロセスに簡略化する方法があります。しかし、クマの生息地は半連続的な性質を持ち、質も多様であるため、これらの仮定には適合しません。
簡略化されたモデリングアプローチの可能性について
最も単純な現象論的アプローチは、襲撃が許容できる水準に達するまでクマを狩猟することです。狩猟は、最も深刻な遭遇が発生する可能性が高い場所、すなわちクマの目撃や襲撃が最も多い地域で実施すべきです。それでもなお、分散個体数を減らすためには、中核地域の個体群密度を低下させる必要があります。また、クマの個体数調査データと襲撃件数の歴史的データを用いると、生態学的・政治的に許容可能な襲撃件数、さらにはクマの個体群規模を設定することもできます。
第二の現象論的アプローチでは、人間とクマの密度の詳細なモデリングを実施することができます。人間の密度は、例えば携帯電話の位置情報データから推定可能であり、そのようなデータが(匿名化された形で)通信事業者から入手できれば問題ありません。クマの餌となる種の分布図や移動に適した生息地の地図は、クマの移動経路を理解するうえで極めて重要です。論理的には、クマの個体群は、人間が少なく、資源が豊富で孤立した地域から、人間が多く、生息地の分断化が進み、資源が乏しい地域へと拡大していきます。
クマとの遭遇を減らすためには、クマの個体数削減(駆除・狩猟)か、クマの多い地域への人の訪問の削減、あるいはその両方が必要となります。例えば、子連れのメスが多い地域を避けるなど、最悪の遭遇を減らす余地はあるかもしれません。クマの個体群の自然増加率が非常に高いため、駆除なしにクマによる襲撃を制御下に置くことは非現実的であると思われます。
第三の現象論的アプローチの選択肢は、空間的個体群動態の完全なモデルです。このようなモデルに関連する可能性がある要因には、年齢別の繁殖率と死亡率、採餌や繁殖に関連する資源密度、移動や分散に対する生息地の選好性と適性、種固有の移動速度と距離、繁殖・生存・移動に対する密度依存性の影響、年間の気象条件が及ぼす影響、主要な捕食対象種や競合種との種間相互作用、初期個体群状態、人間との相互作用などが含まれます。これらをすべて含めようとすると、統一的なパラメータ化が困難になり、パラメータ間の相関関係を見逃した場合など、予測を誤る可能性もあります。簡略化策として、構造化個体群モデルに簡略化された空間的要素を組み合わせ、クマ個体群の分布を近似する手法が考えられます。
クマの個体群のモデリングと順応的管理には、襲撃発生地での狩猟から本格的な空間個体制御モデルまで、幅広い選択肢が存在します。これらの手法は、要件、複雑さ、信頼性、実装速度、適用可能性において大きく異なります。より単純な手法から始めることによって、過剰なクマの襲撃への初期対応をより迅速におこなうことが可能となります。


